Базальный (филогенетика) - Basal (phylogenetics) - Wikipedia
В филогенетика, базальный это направление основание (или корень) укорененное филогенетическое дерево или же кладограмма. Термин может более строго применяться только к узлам, смежным с корнем, или более свободно к узлам, рассматриваемым как близкие к корню. Каждому узлу в дереве соответствует клады; т.е. клад C может быть описан как базальный в более крупной кладе D если его корень напрямую связан с корнем D. Условия глубоко ветвящийся или же раннее ветвление похожи по смыслу.
Хотя всегда должно быть две или более одинаковых базальных клады, прорастающих из корня каждой кладограммы, эти клады могут сильно различаться по таксономический ранг,[n 1] видовое разнообразие, или оба.[n 2] Если C базальная клада внутри D который имеет самый низкий ранг среди всех базальных клад в пределах D,[n 3] C можно описать как в базальный таксон этого ранга в D.[n 4] Концепция "ключевые инновации 'подразумевает некоторую степень корреляции между эволюционными инновациями и диверсификация.[1][2][3][n 5] Однако такая корреляция не делает данный случай предсказуемым, поэтому наследственные персонажи не следует приписывать членам базальной клады с меньшим видовым разнообразием без дополнительных доказательств.[4][5][6][n 6]
В общем кладе А базальнее клады B если B является подгруппой сестринская группа из А или из А сам.[n 7] Внутри больших групп термин «базальный» может использоваться в широком смысле для обозначения «ближе к корню, чем подавляющее большинство», и в этом контексте может возникнуть такая терминология, как «очень базальный». «Основная клада» - это клада, представляющая все, кроме базальной клады (ки) самого низкого ранга в пределах более крупной клады; например., ядро эвдикотов.
использование
Базальная группа в более строгом смысле образует сестринскую группу по отношению к остальной части более крупной клады, как в следующем случае:
| Базальная клада # 2 |
Хотя на такой кладограмме легко определить базальную кладу, уместность такой идентификации зависит от точности и полноты диаграммы. В этом примере предполагается, что концевые ветви кладограммы изображают все сохранившиеся таксоны данного ранга внутри клады; в противном случае диаграмма может оказаться весьма обманчивой. Кроме того, эта квалификация не гарантирует, что представлено разнообразие вымерших таксонов (которые могут быть плохо изучены).
В филогенетике термин базальный может быть объективно применен к кладами организмов, но, как правило, применяется избирательно и более спорно к группам или родословная[n 8] считалось, что они обладают наследственными признаками, или таким предполагаемым наследственным черты самих себя. При описании персонажей "предки" или "плезиоморфный "предпочтительнее" базального "или"примитивный ", последнее из которых может иметь ложный оттенок неполноценности или отсутствия сложности.
Несмотря на повсеместное использование базальный, немного систематики считают, что его применение к существующим группам не нужно и вводит в заблуждение.[9] Этот термин чаще применяется, когда одна ветвь (та, которая считается «базальной») менее разнообразна, чем другая ветвь (это ситуация, в которой можно было бы ожидать найти базальный таксон более низкого минимум классифицировать). Этот термин может быть двусмысленным, поскольку он также относится к направлению корня дерева, которое представляет гипотетического предка; следовательно, это может неточно означать, что сестринская группа более богатой видами клады проявляет наследственные черты.[6] Сохранившаяся базальная группа может напоминать, а может и не напоминать последний общий предок из более крупной клады в большей степени, чем другие группы, и отделен от этого предка на такое же количество времени, что и все остальные существующие группы. Однако бывают случаи, когда необычно маленький размер сестринской группы действительно коррелирует с необычным количеством наследственных черт, как в Амборелла (Смотри ниже). Другими известными примерами этого явления являются яйцекладущий размножение и без сосков кормление грудью из монотремы, базальная клада млекопитающих[10] всего с пятью видами, а архаичная анатомия Tuatara,[11] базальная клада лепидозавр с одним видом.
Примеры
Цветущие растения

В цветущее растение семья Amborellaceae, ограниченный Новая Каледония в юго-западной части Тихого океана,[n 9] это базальная клада современных покрытосеменных растений,[12] состоящий из большинства основных видов, родов, семейств и отрядов внутри группы (из всего около 250 000 видов покрытосеменных). Черты Amborella trichopoda считаются полезными для понимания эволюции цветковых растений; например, у него «самая примитивная древесина (состоящая только из трахеиды ) любого живого покрытосеменного », а также« простых отдельных частей цветка с неопределенным числом и незапечатанных плодолистиков ».[13] Однако эти черты представляют собой смесь архаики и апоморфный (производные) признаки, которые были отсортированы только путем сравнения с другими покрытосеменными, и их положения в филогенетическом дереве ( Окаменелости потенциально может быть полезным в этом отношении, но отсутствует в данном случае).[13] Кладограмма ниже основана на Рамирес-Барахоне. и другие. (2020),[14] с подсчетом видов, взятым из указанного источника.
|
Человекообразные обезьяны
Среди человекообразных обезьян гориллы (восточные и западные) являются родственной группой обычных шимпанзе, бонобо и людей. Эти пять видов образуют кладу, подсемейство Гоминины (Африканские обезьяны), из которых Горилла базальный род. Однако, если анализ не ограничивается родами, Гомо плюс Сковорода клады также базальные.

| Гоминины |
| ||||||||||||||||||||||||
Более того, орангутаны являются сестринской группой по отношению к Homininae и являются базальным родом в семействе больших обезьян. Гоминиды в целом.
| Гоминиды |
| ||||||||||||||||||
Подсемейства Homininae и Ponginae оба являются базальными в пределах Hominidae, но, учитывая, что на кладограмме нет небазальных подсемейств, маловероятно, что этот термин будет применяться к обоим. В общем, утверждение о том, что одна группа (например, орангутанги) является основной или ответвляется первой внутри другой группы (например, Hominidae), может не иметь смысла, если только соответствующий таксономический уровень (уровни) (род, в данном случае ) указан. Если этот уровень не может быть указан (т. Е. Если рассматриваемая клада не имеет ранга), может потребоваться более подробное описание соответствующих сестринских групп.
В этом примере орангутаны отличаются от других родов своим азиатским ареалом. Этот факт в сочетании с их базовым статусом указывает на то, что самый недавний общий предок современных человекообразных обезьян мог быть евразийцем (см. Ниже), что согласуется с другими доказательствами.[18] (Конечно, меньшие обезьяны полностью азиатские.) Орангутаны также отличаются от африканских обезьян более высокой степенью древесный образ жизни - черта, которую обезьяны считали наследственной.[19][20]
Актуальность для биогеографической истории
Учитывая, что наиболее глубокий филогенетический раскол в группе, вероятно, произошел в начале ее истории, идентификация наиболее базальных субкладов в широко рассредоточенном таксоне или кладе может дать ценную информацию о регионе ее происхождения. В некоторых ситуациях, когда это может быть неочевидно, также можно сделать вывод о направлении миграции от зоны происхождения (как в Amaurobioides и случаи Noctilionoidea ниже). Примеры включают:
- Пауки из рода Amaurobioides присутствуют в Южной Африке, Австралии, Новой Зеландии и Чили.[21][22] Самая базальная клада - южноафриканская; Свидетельства последовательности ДНК показывают, что после того, как их южноамериканские предки достигли Южной Африки, они рассредоточены на восток вплоть до Южной Америки с интервалом около 8 миллионов лет.[22]
- Игуанид ящерицы (Sensu lato ) распространяются по всей Америке, на Мадагаскар, и дальше Фиджи и Тонга в западной части южной части Тихого океана. В Малагасийские формы являются базальными, с предполагаемой датой расхождения с другими, составляющей ~ 162 миллиона лет, незадолго до момента отделения Мадагаскара от Африки.[23] Это говорит о том, что игуаниды когда-то имели широкое распространение. Гондванец распределение; после того, как представители Малагасийского и Нового Света были разделены викарианс, менее изолированные игуаниды Старого Света вымерли из-за конкуренции с другими группами ящериц (например, агамиды ). В отличие, игуаниды западной части Тихого океана глубоко гнездятся в американских игуанидах,[24] очевидно колонизировав свой изолированный ареал после эпических 10000 км рафтинг.[25][26]
- Коралловые змеи составляют около 16 видов в Азии и более 65 видов в Северной и Южной Америке. Однако ни один из американских кладов не является базальным, что подразумевает, что родословная группы происходила из Старого Света.[27]
- Современные австралидельфские сумчатые составляют около 240 видов в Австралазии и один вид ( Монито дель Монте ) В Южной Америке. Тот факт, что monito del monte занимает базальное положение (самый базальный вид, род, семейство и отряд) в надотряде Австралиделфия - важный ключ к разгадке того, что он произошел в Южной Америке. Этот вывод согласуется с тем, что южноамериканский порядок Дидельфиморфия базальна в инфраклассе Marsupialia; То есть, современные сумчатые в целом также, по-видимому, произошли из Южной Америки.[17][n 10][n 11][n 12]
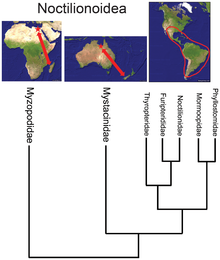
- Пока надсемейство летучих мышей Noctilionoidea насчитывает более 200 видов в Неотропах, два в Новой Зеландии, и двое на Мадагаскаре, базальное положение малагасийской семьи[34] предполагает, в сочетании с летописью окаменелостей и следующим по базальному уровню расположением новозеландской семьи, что суперсемейство возникло в Африке, а затем мигрировало на восток в Южную Америку, размножаясь там, но выживая в Старом Свете только в Refugia.[35]
- Род Уроцион (серая и островная лисы) базальна в подсемейство собак,[36] предполагая североамериканское происхождение почти всемирной группы. Это согласуется с данными окаменелостей, указывающими на североамериканское происхождение семья псовых в целом (два других подсемейства псовых, вымершие Borophaginae[37] и Hesperocyoninae,[38] последние были базальными у Canidae, оба эндемичный в Северную Америку).
Примечания
- ^ Означает самые низкие таксономические ранги соответствующих клад; высшие ранги должны быть одинаковыми (при условии, что они ранжированы).
- ^ Увидеть Амборелла пример, в котором одна базальная клада является единственной сохранившейся разновидность (это также единственный живой представитель порядок, Amborellales ). Между тем, другая (не ранжированная) сестринская базальная клада насчитывает около 250 000 видов.
- ^ Например, C может быть род а другая базальная клада (и) может иметь более высокие ранги подсемейство или же семья.
- ^ в пример великих обезьян, Горилла - базальный род подсемейства Homininae, а Понго является базальным родом семейства Hominidae. Две базальные клады последнего имеют наивысший ранг подсемейства, то есть Homininae и Ponginae.
- ^ Большая диверсификация клады также может быть связана с колонизацией нового массива суши, особенно если он больше или менее конкурентоспособен, чем массив земли предков; увидеть примеры коралловых змей, сумчатых и ноктилионоидных летучих мышей.
- ^ Например, гигантская панда представляет собой наиболее базальный из существующих видов, род и подсемейство в пределах Ursidae,[7] но его специализация в бамбуковой диете - не наследственные персонажи-медвежата.[8]
- ^ То есть на диаграмме ниже обе базальные клады # 1 и # 2 более базальные, чем небазальные клады # 1, которая, в свою очередь, более базальна, чем небазальные клады # 2 и # 3.
- ^ Поскольку родословная - это линейная цепочка потомков, все родословные внутри клады можно проследить не только до корня, но и до происхождение жизни. Таким образом, с филогенетической точки зрения представление о родословной как о базальной бессмысленно. Однако в генетике базальная линия относится к линии, соединяющей общего предка с одним вариантом аллель к предку ветви с двумя вариантами потомка.
- ^ Новая Каледония рассматривается как рефугиум; то есть в этом случае считается, что географическое положение базальной клады не является свидетельством того места происхождения покрытосеменных.
- ^ Эти выводы подтверждаются открытием эоцен ископаемые останки микробиотериан Вудбурнодон казеи в Антарктида,[28] который, как предполагается, служил промежуточной станцией на пути миграции в Австралию до окончательного распада Гондвана.
- ^ Точно так же среди австралобатрахийских лягушек южноамериканское семейство Калиптоцефалеллиды, с 5 существующими видами (живущими в той же Вальдивский лес как Монито дель Монте), является базовым для австралийских семей Limnodynastidae и Myobatrachidae,[29] насчитывает около 120 современных видов, что позволяет предположить южноамериканское происхождение группы.[30] Это согласуется с находкой окаменелости южноамериканской семьи в Антарктиде.[31]
- ^ Ratites возможно, аналогичным образом путешествовали по суше из Южной Америки, чтобы колонизировать Австралию;[32] ископаемый ратит известен из Антарктиды,[33] и южноамериканские реи более базальные в группе, чем австрало-тихоокеанские ратиты.[32]
Рекомендации
- ^ Heard, S.B .; Хаузер, Д. (1995). «Ключевые эволюционные инновации и их экологические механизмы». Историческая биология. 10 (2): 151–173. Дои:10.1080/10292389509380518.
- ^ Энгель, M.S .; Гримальди, Д.А. (2004). «Новый свет пролил свет на древнейшее насекомое». Природа. 427 (6975): 627–630. Дои:10.1038 / природа02291. PMID 14961119. S2CID 4431205.
- ^ Fernández ‐ Mazuecos, M .; Blanco ‐ Pastor, J.L .; Хуан, А .; Carnicero, P .; Форрест, А .; Alarcón, M .; Vargas, P .; Гловер, Б.Дж. (2019). «Макроэволюционная динамика нектарных шпор, ключевое эволюционное новшество». Новый Фитолог. 222 (2): 1123–1138. Дои:10.1111 / nph.15654. HDL:10045/89954. PMID 30570752.
- ^ Баум, Д. А. (4 ноября 2013 г.). «Филогенетика и история жизни». Принстонский путеводитель по эволюции. Издательство Принстонского университета. п. 57. ISBN 978-1-4008-4806-5. OCLC 861200134.
- ^ Crisp, M.D .; Кук, Л. Г. (март 2005 г.). «Означают ли ранние ветвящиеся линии наследственные черты?». Тенденции в экологии и эволюции. 20 (3): 122–128. Дои:10.1016 / j.tree.2004.11.010. PMID 16701355.
- ^ а б Дженнер, Рональд А (2006). «Разгрузка эво-дево: исконные достопримечательности, модельные организмы и базальная чепуха». Гены развития и эволюция. 216 (7–8): 385–394. Дои:10.1007 / s00427-006-0084-5. PMID 16733736. S2CID 23140594.
- ^ Krause, J .; Унгер, Т .; Noçon, A .; Маласпинас, А .; Kolokotronis, S .; Стиллер, М .; Сойбелзон, Л .; Spriggs, H .; Уважаемый, P.H .; Briggs, A.W .; Bray, S.C.E .; О'Брайен, С. Дж .; Rabeder, G .; Matheus, P .; Купер, А .; Слаткин, М .; Pääbo, S .; Хофрейтер, М. (2008). «Митохондриальные геномы показывают взрывное излучение вымерших и современных медведей вблизи границы миоцена и плиоцена». BMC Эволюционная биология. 8 (220): 220. Дои:10.1186/1471-2148-8-220. ЧВК 2518930. PMID 18662376.
- ^ Маклеллан, Брюс; Райнер, Дэвид С. (1994). «Обзор эволюции медведя». Медведи: их биология и менеджмент. 9 (1): 85–96. Дои:10.2307/3872687. JSTOR 3872687.
- ^ Krell, Frank T .; Крэнстон, Питер С. (2004). «Какая сторона дерева прикорневая?». Систематическая энтомология. 29 (3): 279–281. Дои:10.1111 / j.0307-6970.2004.00262.x.
- ^ Ван Риде, Теун (2005). «Утконос на своем месте: ядерные гены и инделы подтверждают родственные связи монотрем и терианцев». Молекулярная биология и эволюция. 23 (3): 587–597. Дои:10.1093 / molbev / msj064. PMID 16291999.
- ^ Эррера-Флорес, Дж. А .; Stubbs, T. L .; Бентон, М. Дж .; Рута, М. (2017). "Макроэволюционные закономерности в Rhynchocephalia: туатара (Sphenodon punctatus) живое ископаемое? ". Палеонтология. 60 (3): 319–328. Дои:10.1111 / pala.12284.
- ^ Li, H.-T .; Yi, T.-S .; Gao, L.-M .; Ma, P.-F .; Zhang, T .; Yang, J.-B .; Gitzendanner, M.A .; Fritsch, P.W .; Cai, J .; Luo, Y .; Wang, H .; van der Bank, M .; Zhang, S.-D .; Wang, Q.-F .; Wang, J .; Zhang, Z.-R .; Fu, C.-N .; Yang, J .; Hollingsworth, P.M .; Чейз, M.W .; Soltis, D.E .; Soltis, P.S .; Ли, Д.-З. (2019). «Происхождение покрытосеменных и загадка юрского промежутка». Природа Растения. 5 (5): 461–470. Дои:10.1038 / s41477-019-0421-0. PMID 31061536. S2CID 146118264.
- ^ а б Эссиг, Ф. Б. (01.07.2014). "Что такого примитивного в Амборелла?". Профессор ботаники. Получено 2014-10-04.
- ^ Ramírez-Barahona, S .; Sauquet, H .; Магальон, С. (2020). «Отсроченная и географически неоднородная диверсификация семейств цветковых растений». Природа Экология и эволюция. 4 (9): 1232–1238. Дои:10.1038 / с41559-020-1241-3. PMID 32632260. S2CID 220375828.
- ^ а б c d е Palmer, J.D .; Soltis, D.E .; Чейз, М.В. (2004). «Растение древо жизни: обзор и некоторые точки зрения». Американский журнал ботаники. 91 (10): 1437–1445. Дои:10.3732 / ajb.91.10.1437. PMID 21652302.
- ^ а б c Christenhusz, M. J. M .; Бинг, Дж. У. (2016). «Число известных видов растений в мире и его ежегодный прирост». Фитотакса. 261 (3): 201–217. Дои:10.11646 / phytotaxa.261.3.1.
- ^ а б Nilsson, M. A .; Чураков, Г .; Sommer, M .; Van Tran, N .; Земанн, А .; Brosius, J .; Шмитц, Дж. (27 июля 2010 г.). Пенни, Дэвид (ред.). «Отслеживание эволюции сумчатых животных с использованием вставок архаичных геномных ретропозонов». PLOS Биология. 8 (7): e1000436. Дои:10.1371 / journal.pbio.1000436. ЧВК 2910653. PMID 20668664.
- ^ Моя-Сола, С .; Альба, Д. М .; Almecija, S .; Казановас-Вилар, I .; Kohler, M .; De Esteban-Trivigno, S .; Robles, J.M .; Галиндо, Дж .; Фортуни, Дж. (16.06.2009). «Уникальный европейский гоминоид среднего миоцена и происхождение клады больших обезьян и людей». PNAS. 106 (24): 9601–9606. Дои:10.1073 / pnas.0811730106. ЧВК 2701031. PMID 19487676..
- ^ MaClatchy, L .; Гебо, Д .; Kityo, R .; Пилбим, Д. (2000). «Посткраниальная функциональная морфология Моротопитекус бишопи, с последствиями для эволюции передвижения современных обезьян ". Журнал эволюции человека. 39 (2): 159–183. Дои:10.1006 / jhev.2000.0407. PMID 10968927.
- ^ Thorpe, S.K.S .; Кромптон, Р. Х. (2006). «Позиционное поведение орангутана и природа древесного передвижения у Hominoidea». Американский журнал физической антропологии. 131 (3): 384–401. Дои:10.1002 / ajpa.20422. PMID 16617429.
- ^ Куксо, Ф. (8 ноября 2016 г.). «Пауки-мореплаватели обошли весь мир за 8 миллионов лет». Scientific American. Получено 2016-11-10.
- ^ а б Кунтнер, М .; Чеккарелли, Ф. С .; Opell, B.D .; Haddad, C. R .; Рэйвен, Роберт Дж .; Сото, Э. М .; Рамирес, М. Дж. (12 октября 2016 г.). «Вокруг света за восемь миллионов лет: историческая биогеография и эволюция пауков зоны опрыскивания. Amaurobioides (Araneae: Anyphaenidae) ". PLOS ONE. 11 (10): e0163740. Дои:10.1371 / journal.pone.0163740. ЧВК 5061358. PMID 27732621.
- ^ Okajima, Y .; Кумазава, Ю. (15.07.2009). "Митогеномные перспективы филогении и биогеографии игуанид: Гондванский викариант по происхождению мадагаскарских oplurines". Ген. 441 (1–2): 28–35. Дои:10.1016 / j.gene.2008.06.011. PMID 18598742.
- ^ Schulte, J. A .; Valladares, J. P .; Ларсон, А. (сентябрь 2003 г.). «Филогенетические отношения внутри игуанид, выведенные с использованием молекулярных и морфологических данных и филогенетической таксономии игуанских ящериц». Herpetologica. 59 (3): 399–419. Дои:10.1655/02-48. S2CID 56054202.
- ^ Гиббонс, Дж. Р. Х. (1981-07-31). "Биогеография Брахилофус (Iguanidae), включая описание нового вида, Б. vitiensis, из Фиджи ". Журнал герпетологии. 15 (3): 255–273. Дои:10.2307/1563429. JSTOR 1563429.
- ^ Keogh, J. S .; Эдвардс, Д. Л; Фишер, Р. Н; Харлоу, П. С. (2008). «Молекулярный и морфологический анализ находящихся под угрозой исчезновения фиджийских игуан показывает загадочное разнообразие и сложную биогеографическую историю». Философские труды Королевского общества B: биологические науки. 363 (1508): 3413–3426. Дои:10.1098 / rstb.2008.0120. ЧВК 2607380. PMID 18782726.
- ^ Словински, Дж. Б.; Boundy, J .; Лоусон, Р. (июнь 2001 г.). «Филогенетические отношения азиатских коралловых змей (Elapidae: Calliophis и Maticora) на основе морфологических и молекулярных признаков». Herpetologica. 57 (2): 233–245. JSTOR 3893186.
- ^ Goin, F.J .; Zimicz, N .; Регуэро, М. А .; Santillana, S.N .; Marenssi, S.A .; Моли, Дж. Дж. (2007). «Новое сумчатое животное (Mammalia) из эоцена Антарктиды, а также происхождение и родство Microbiotheria». Revista de la Asociación Geológica Argentina. 62 (4): 597–603. Получено 2016-07-17.
- ^ Pyron, R.A .; Винс, Дж. Дж. (2011). «Крупномасштабная филогения амфибий, включающая более 2800 видов, и пересмотренная классификация существующих лягушек, саламандр и цецилий». Молекулярная филогенетика и эволюция. 61 (2): 543–583. Дои:10.1016 / j.ympev.2011.06.012. PMID 21723399.
- ^ Feng, Y.-J .; Блэкберн, округ Колумбия; Liang, D .; Hillis, D.M .; Wake, D.B .; Каннателла, округ Колумбия; Чжан, П. (2017). «Филогеномика показывает быструю одновременную диверсификацию трех основных клад гондванских лягушек на границе мела и палеогена». Труды Национальной академии наук. 114 (29): E5864 – E5870. Дои:10.1073 / pnas.1704632114. ЧВК 5530686. PMID 28673970.
- ^ Mörs, T .; Reguero, M .; Василян, Д. (2020). «Первая ископаемая лягушка из Антарктиды: последствия для климатических условий высоких широт в эоцене и гондванского космополитизма в Австралии». Научные отчеты. 10 (1). Дои:10.1038 / s41598-020-61973-5. PMID 32327670.
- ^ а б Yonezawa, T .; Сегава, Т .; Mori, H .; Кампос, П. Ф .; Hongoh, Y .; Endo, H .; Акиёси, А .; Коно, Н .; Nishida, S .; Wu, J .; Jin, H .; Adachi, J .; Кишино, Х .; Курокава, К .; Nogi, Y .; Tanabe, H .; Мукояма, H .; Yoshida, K .; Rasoamiaramanana, A .; Yamagishi, S .; Hayashi, Y .; Yoshida, A .; Koike, H .; Акишинономия, Ф .; Willerslev, E .; Хасегава, М. (15 декабря 2016 г.). «Филогеномика и морфология вымерших палеогнатов раскрывают происхождение и эволюцию Ratites». Текущая биология. 27 (1): 68–77. Дои:10.1016 / j.cub.2016.10.029. PMID 27989673.
- ^ Tambussi, C.P .; Noriega, J.I .; Gazdzicki, A .; Татур, А .; Reguero, M.A .; Бискайно, С.Ф. (1994). "Птица-ратит из палеогеновой формации Ла-Месета, остров Сеймур, Антарктида" (PDF). Польские полярные исследования. 15 (1–2): 15–20. Получено 28 декабря 2019.
- ^ Тилинг, E.C .; Springer, M .; Madsen, O .; Bates, P .; O'Brien, S .; Мерфи, У. (28 января 2005 г.). «Молекулярная филогения летучих мышей освещает биогеографию и летопись окаменелостей». Наука. 307 (5709): 580–584. Дои:10.1126 / science.1105113. PMID 15681385. S2CID 25912333.
- ^ Gunnell, G.F .; Simmons, N.B .; Зайфферт, Э. Р. (04.02.2014). «Новые Myzopodidae (Chiroptera) из позднего палеогена Египта: исправленный семейный диагноз и биогеографические истоки Noctilionoidea». PLoS ONE. 9 (2): e86712. Дои:10.1371 / journal.pone.0086712. ЧВК 3913578. PMID 24504061.
- ^ Lindblad-Toh, K .; Wade, C.M .; Mikkelsen, T.S .; Карлссон, E.K .; Jaffe, D.B .; Kamal, M .; Зажим, М .; Chang, J.L .; Kulbokas, E.J .; Zody, M.C .; Mauceli, E .; Се, X .; Breen, M .; Уэйн, Р.К .; Ostrander, E.A .; Ponting, C.P .; Галиберт, Ф .; Smith, D.R .; Dejong, P.J .; Киркнесс, Э .; Alvarez, P .; Biagi, T .; Brockman, W .; Батлер, Дж .; Chin, C.-W .; Кук, А .; Cuff, J .; Daly, M.J .; Decaprio, D .; и другие. (2005). «Последовательность генома, сравнительный анализ и структура гаплотипов домашней собаки». Природа. 438 (7069): 803–819. Bibcode:2005Натура.438..803л. Дои:10.1038 / природа04338. PMID 16341006.
- ^ Ван, Сяомин; Тедфорд, Ричард; Тейлор, Берил (1999-11-17). «Филогенетическая систематика Borophaginae (Carnivora, Canidae)». Бюллетень Американского музея естественной истории. 243. HDL:2246/1588.
- ^ Ван, X .; Тедфорд, Р. Х. (2008). Собаки, их ископаемые родственники и эволюционная история. Издательство Колумбийского университета. С. 23–31. ISBN 978-0-231-13528-3.
внешняя ссылка
- «Интерпретация древовидной диаграммы или списка подгрупп на странице« Древо жизни »». Веб-проект "Древо жизни".
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||