Наследственность - Heritability

Наследственность это статистика используется в полях разведение и генетика что оценивает степень вариация в фенотипический признак в Население это связано с генетическая вариация между людьми в этой популяции.[1] Он измеряет, насколько вариативность признака может быть связана с вариацией генетических факторов, а не с вариацией факторов окружающей среды. Концепция наследственности может быть выражена в форме следующего вопроса: «Какова доля вариаций данного признака в популяции, которая не объясняется средой или случайностью? "[2]
Другие причины измеряемой вариации признака характеризуются как экологические факторы, в том числе ошибка наблюдения. В исследованиях наследственности на людях они часто делятся на факторы, относящиеся к «общей среде» и «неразделенной среде», в зависимости от того, приводят ли они к тому, что люди, выросшие в одном доме, более или менее похожи на людей, которые не были такими.
Наследственность оценивается путем сравнения индивидуальных фенотипических вариаций среди родственных особей в популяции, путем изучения связи между индивидуальным фенотипом и данными генотипа.[3][4] или даже путем моделирования сводных данных из исследования ассоциации всего генома (GWAS).[5] Наследственность - важное понятие в количественная генетика, особенно в селекция и генетика поведения (например, исследования близнецов ). Это источник большой путаницы из-за того, что его техническое определение отличается от его общепринятого народного определения. Следовательно, его использование создает неправильное впечатление, что поведенческие черты «наследуются» или специально передаются через гены.[6] Поведенческие генетики также проводят анализ наследственности, исходя из предположения, что гены и окружающая среда вносят отдельный дополнительный вклад в поведенческие черты.[7]
Обзор
Наследственность измеряет долю изменчивости фенотипа, которая может быть отнесена к генетическая вариация. Это не то же самое, что сказать, что эта часть индивидуального фенотипа обусловлена генетикой. Например, неверно утверждать, что, поскольку наследуемость черт личности составляет около 0,6, это означает, что 60% вашей личности унаследовано от родителей, а 40% - из окружающей среды. Кроме того, наследственность может измениться без каких-либо генетических изменений, например, когда среда начинает вносить вклад в большее разнообразие. В качестве примера рассмотрим, что оба гены и окружающая среда могут влиять на интеллект. Наследственность может увеличиться, если увеличится генетическая изменчивость, в результате чего люди будут демонстрировать больше фенотипических вариаций, например, показывать разные уровни интеллекта. С другой стороны, наследственность может также увеличиться, если изменчивость окружающей среды уменьшается, в результате чего люди демонстрируют меньшие фенотипические вариации, например, демонстрируют более схожие уровни интеллекта. Наследственность увеличивается, когда генетические факторы вносят больший вклад в изменение или потому, что негенетические факторы вносят вклад в меньшую изменчивость; важен относительный вклад. Наследственность специфична для определенной популяции в определенной среде. Следовательно, высокая наследуемость признака не обязательно означает, что признак не очень подвержен влиянию окружающей среды.[8] Наследственность также может измениться в результате изменений среды, миграции, инбридинг, или способ измерения самой наследственности в исследуемой популяции.[9] Наследственность признака не следует интерпретировать как меру того, в какой степени указанный признак генетически детерминирован у человека.[10][11]
Степень зависимости фенотипа от окружающей среды также может быть функцией задействованных генов. Вопросы наследственности сложны, потому что гены могут канализировать фенотип, что делает его проявление почти неизбежным во всех средах. Люди с одним и тем же генотипом могут также проявлять разные фенотипы с помощью механизма, называемого фенотипическая пластичность, что в некоторых случаях затрудняет измерение наследственности. Недавние исследования в области молекулярной биологии выявили изменения в транскрипционный активность отдельных генов, связанных с изменениями окружающей среды. Однако существует большое количество генов, транскрипция которых не зависит от окружающей среды.[12]
Оценки использования наследственности Статистический анализ чтобы помочь выявить причины различий между людьми. Поскольку наследственность связана с изменчивостью, она обязательно связана с различиями между людьми в популяции. Наследственность может быть одномерный - изучение одного признака - или многовариантное - изучение генетических и экологических ассоциаций между несколькими признаками одновременно. Это позволяет проверить генетическое совпадение между различными фенотипами: например, цвет волос и цвет глаз. Окружающая среда и генетика также могут взаимодействовать, и анализ наследственности может проверять и изучать эти взаимодействия (модели GxE).
Предпосылкой для анализа наследуемости является наличие некоторой изменчивости популяции, которую необходимо учитывать. Этот последний пункт подчеркивает тот факт, что наследуемость не может учитывать влияние факторов, которые инвариантны в популяции. Факторы могут быть инвариантными, если они отсутствуют и не существуют в популяции, например, никто не имеет доступа к определенному антибиотик, или потому что они вездесущи, например, если все пьют кофе. На практике все поведенческие черты человека различаются, и почти все черты проявляют некоторую наследственность.[13]
Определение
Любой конкретный фенотип может быть смоделированный как сумма генетических и экологических эффектов:[14]
- Фенотип (п) = Генотип (г) + Окружающая среда (E).
Аналогичным образом фенотипическая дисперсия признака - Var (P) - представляет собой сумму следующих эффектов:
- Вар (п) = Вар (г) + Var (E) + 2 Cov (г,E).
В запланированном эксперименте Cov (г,E) можно контролировать и удерживать на уровне 0. В этом случае наследуемость, определяется как:[15]


ЧАС2 наследственность в широком смысле. Это отражает весь генетический вклад в фенотипическую изменчивость популяции, включая аддитивные, доминирующий, и эпистатический (мультигенные взаимодействия), а также материнские и отцовские эффекты, где люди напрямую зависят от фенотипа своих родителей, например, с молоко продукция у млекопитающих.
Особенно важным компонентом генетической дисперсии является аддитивная дисперсия Var (A), которая представляет собой дисперсию, обусловленную средними эффектами (аддитивными эффектами) аллели. Поскольку каждый родитель передает один аллель на локус для каждого потомства сходство родителей и потомков зависит от среднего эффекта отдельных аллелей. Следовательно, аддитивная дисперсия представляет собой генетический компонент дисперсии, ответственный за сходство между родителями и потомками. Аддитивная генетическая часть фенотипической дисперсии известна как узко-смысловая наследственность и определяется как

Верхний регистр ЧАС2 используется для обозначения широкого смысла, а нижний регистр час2 в узком смысле.
Для признаков, которые не являются непрерывными, но дихотомическими, такими как дополнительный палец ноги или определенные заболевания, вклад различных аллелей можно рассматривать как сумму, которая после порогового значения проявляется как признак, дающий модель порога ответственности в котором можно оценить наследуемость и смоделировать выбор.
Добавочная дисперсия важна для отбор. Если оказывается давление отбора, такое как улучшение поголовья, реакция признака напрямую связана с узкой наследуемостью. Среднее значение признака будет увеличиваться в следующем поколении в зависимости от того, насколько среднее значение выбранных родителей отличается от среднего значения популяции, из которой были выбраны выбранные родители. Наблюдаемые ответ на выбор приводит к оценке наследуемости в узком смысле (называемой реализованная наследственность). Это принцип, лежащий в основе искусственный отбор или разведение.
пример
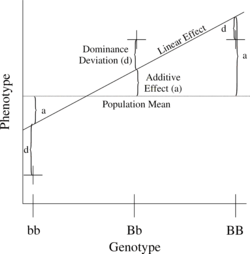
Простейшая генетическая модель включает один локус с двумя аллелями (b и B), влияющими на один количественный фенотип.
Номер B аллели могут варьироваться от 0, 1 или 2. Для любого генотипа BяBj, ожидаемый фенотип может быть записан как сумма общего среднего, линейного эффекта и отклонения доминирования:
- = Средняя численность населения + Аддитивный эффект () + Отклонение доминирования ().



Аддитивная генетическая дисперсия в этом локусе - это средневзвешенное квадратов аддитивных эффектов:

где

Аналогичная зависимость существует и для дисперсии отклонений доминирования:

где

В линейная регрессия Соотношение фенотипа к генотипу показано на рисунке 1.
Предположения
Оценки общей наследуемости человеческих черт предполагают отсутствие эпистаза, что получило название «допущение аддитивности». Хотя некоторые исследователи цитируют такие оценки в поддержку существования "отсутствие наследственности «не учитываемые известными генетическими локусами, предположение об аддитивности может сделать эти оценки недействительными.[16] Есть также некоторые эмпирические доказательства того, что предположение об аддитивности часто нарушается в поведенческих генетических исследованиях интеллекта и академическая успеваемость.[17]
Оценка наследственности
Поскольку только п могут наблюдаться или измеряться напрямую, наследуемость должна оцениваться по сходству, наблюдаемому у субъектов, различающихся по уровню генетического или средового сходства. В статистический анализы, необходимые для оценки генетический и экологический компоненты дисперсии зависят от характеристик выборки. Вкратце, лучшие оценки получаются с использованием данных, полученных от людей с сильно различающимися уровнями генетического родства, такими как двойняшки, братья и сестры, родители и потомки, а не от более отдаленных (и, следовательно, менее похожих) субъектов. В стандартная ошибка оценки наследственности улучшаются при больших размерах выборки.
В популяциях нечеловеческого происхождения часто можно собирать информацию контролируемым способом. Например, среди сельскохозяйственных животных легко заставить быка производить потомство от большого количества коров и контролировать окружающую среду. Такие экспериментальный контроль как правило, невозможно при сборе данных о людях, полагаясь на естественные отношения и окружающую среду.
В классической количественной генетике существовало две точки зрения относительно оценки наследственности.
Один школа мысли был разработан Сьюэлл Райт в Чикагский университет, и далее популяризируется К. К. Ли (Чикагский университет ) и Дж. Л. Лаш (Государственный университет Айовы ). Он основан на анализе корреляций и, как следствие, регрессии. Анализ пути был разработан Сьюэлл Райт как способ оценки наследственности.
Второй изначально был разработан Р. А. Фишер и расширился на Эдинбургский университет, Государственный университет Айовы, и Университет штата Северная Каролина, а также другие школы. Он основан на дисперсионный анализ селекционных исследований с использованием внутриклассовой корреляции родственников. Различные методы оценки компонентов дисперсии (и, следовательно, наследуемости) от ANOVA используются в этих анализах.
Сегодня о наследственности можно судить по общим родословным, используя линейные смешанные модели и из геномное родство оценивается по генетическим маркерам.
В исследованиях наследственности человека часто используются схемы исследования усыновления, часто с идентичные близнецы которые были разлучены в раннем возрасте и выросли в разных условиях. Такие люди имеют одинаковые генотипы и могут использоваться для разделения эффектов генотипа и окружающей среды. Ограничением такой конструкции является обычная пренатальная среда и относительно небольшое количество близнецов, воспитываемых отдельно. Второй и более распространенный дизайн - это исследование близнецов в котором сходство однояйцевых и разнояйцевых близнецов используется для оценки наследственности. Эти исследования могут быть ограничены тем фактом, что однояйцевые близнецы являются не полностью генетически идентичен, что может привести к недооценке наследственности.
В наблюдательные исследования, или из-за вызывающих воспоминаний эффектов (когда геном вызывает окружающую среду своим воздействием на нее), G и E могут совпадать: корреляция генной среды. В зависимости от методов, используемых для оценки наследуемости, корреляции между генетическими факторами и разделяемой или не разделяемой средой могут или не могут быть смешаны с наследуемостью.[18]
Регрессионные / корреляционные методы оценки
Первая школа оценки использует регрессию и корреляцию для оценки наследуемости.
Сравнение близких родственников
При сравнении родственников мы обнаруживаем, что в целом

где р можно рассматривать как коэффициент родства, б коэффициент регрессии и т - коэффициент корреляции.
Регрессия родитель-потомок

Наследственность может быть оценена путем сравнения родительских и потомственных признаков (как на рис. 2). Наклон линии (0,57) аппроксимирует наследуемость признака, когда значения потомства регрессируют относительно среднего признака у родителей. Если используется значение только одного родителя, то наклон в два раза больше наследуемости. (Обратите внимание, что это источник термина "регресс, "поскольку потомки всегда стремятся вернуться к среднему ценность для населения, т.е., наклон всегда меньше единицы). Этот эффект регрессии также лежит в основе Метод ДеФриса – Фулкера для анализа близнецов, выбранных для одного больного члена.[19]
Сравнение братьев и сестер
Базовый подход к наследуемости может быть применен с использованием моделей полного родства: сравнение сходства между братьями и сестрами, у которых есть как биологическая мать, так и отец.[20] Когда есть только аддитивное действие гена, эта фенотипическая корреляция сиблингов является показателем знакомство - сумма половины аддитивной генетической дисперсии плюс полный эффект общей среды. Таким образом, он устанавливает верхний предел аддитивной наследуемости, вдвое превышающий фенотипическую корреляцию полного Sib. В планах Half-Sib сравниваются фенотипические черты братьев и сестер, у которых один родитель совпадает с другими группами братьев и сестер.
Исследования близнецов
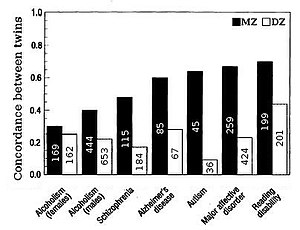
Наследственность признаков у людей чаще всего оценивается путем сравнения сходства между близнецами. «Преимущество близнецовых исследований состоит в том, что общую дисперсию можно разделить на генетические, общие или общие экологические компоненты и уникальные экологические компоненты, что позволяет точно оценить наследуемость».[21] Братские или дизиготные (ДЗ) близнецы в среднем разделяют половину своих генов (при условии, что ассортативная вязка по признаку), поэтому однояйцевые или монозиготные (MZ) близнецы в среднем генетически в два раза более похожи, чем близнецы DZ. Таким образом, грубая оценка наследуемости примерно вдвое превышает разницу в корреляция между близнецами МЗ и ДЗ, т.е. Формула фальконера ЧАС2= 2 (г (MZ) -r (DZ)).
Влияние совместной среды, c2, способствует сходству между братьями и сестрами из-за общности среды, в которой они выросли. Общая среда аппроксимируется корреляцией DZ минус половина наследственности, то есть степень, в которой близнецы DZ имеют одни и те же гены, c2= ДЗ-1/2час2. Уникальное экологическое разнообразие, е2, отражает степень, в которой однояйцевые близнецы, выросшие вместе, не похожи друг на друга, е2= 1-г (MZ).
Анализ дисперсионных методов оценки
Второй набор методов оценки наследуемости включает дисперсионный анализ и оценку компонентов дисперсии.
Базовая модель
Мы используем основное обсуждение Кемпторна.[14] Рассматривая только самые основные генетические модели, мы можем посмотреть на количественный вклад одного локуса в генотип. гя так как

где это влияние генотипа гя и это экологический эффект.


Рассмотрим эксперимент с группой производителей и их потомков от случайных маток. Поскольку потомство получает половину своих генов от отца и половину от (случайной) матери, уравнение потомства имеет вид

Внутриклассовые корреляции
Рассмотрим эксперимент выше. Мы можем сравнить две группы потомства. Первый - это сравнение различного потомства от одного производителя (называемого в группе производителей). Дисперсия будет включать термины для генетической дисперсии (поскольку не все они имеют один и тот же генотип) и дисперсии окружающей среды. Это считается ошибка срок.
Вторая группа потомков - это сравнения средних значений полусибсов друг с другом (называемые среди группы производителей). В добавок к срок ошибки как и во внутригрупповых группах производителей, у нас есть дополнительный термин из-за различий между разными средними показателями полукровок. Внутриклассовая корреляция
- ,

поскольку экологические эффекты не зависят друг от друга.
ANOVA
В эксперименте с производители и потомство на одного производителя, мы можем рассчитать следующий ANOVA, используя как генетическая изменчивость и как экологическая дисперсия:




| Источник | d.f. | Средний квадрат | Ожидаемый средний квадрат |
|---|---|---|---|
| Среди групп производителей | |||
| Внутри групп производителей |






В срок - это внутриклассовая корреляция среди сводных братьев и сестер. Мы легко можем рассчитать . Ожидаемый средний квадрат рассчитывается на основе взаимоотношений особей (например, все потомство в пределах одного производителя являются полукровными братьями) и понимания внутриклассовых корреляций.


Использование ANOVA для расчета наследуемости часто не учитывает наличие взаимодействие генов с окружающей средой, потому что ANOVA имеет гораздо более низкий статистическая мощность для тестирования эффектов взаимодействия, чем для прямых эффектов.[22]
Модель с аддитивными и доминирующими условиями
Для модели с аддитивными членами и членами доминирования, но не с другими, уравнение для одного локуса имеет вид

где
это аддитивный эффект ith аллель - аддитивный эффект jth аллель отклонение доминирования для ijth генотип и это окружающая среда.


Эксперименты можно проводить с настройками, аналогичными приведенным в таблице 1. Используя разные группы взаимосвязей, мы можем оценивать различные внутриклассовые корреляции. С помощью как аддитивная генетическая дисперсия и как дисперсия отклонения доминирования, внутриклассовые корреляции становятся линейные функции этих параметров. В общем,


- Внутриклассовая корреляция

где и находятся как

П[ аллели взятые наугад из пары отношений, идентичны по происхождению ], и

П[ генотипы взятые наугад из пары отношений, идентичны по происхождению ].

Некоторые общие соотношения и их коэффициенты приведены в таблице 2.
| Отношения | ||
|---|---|---|
| Идентичные близнецы | ||
| Родитель-потомство | ||
| Половина братьев и сестер | ||
| Полные братья и сестры | ||
| Двоюродные братья | ||
| Двойные двоюродные братья |






Линейные смешанные модели
В литературе описано большое разнообразие подходов с использованием линейных смешанных моделей. С помощью этих методов фенотипическая дисперсия разделяется на вариации генетической, окружающей среды и экспериментального дизайна для оценки наследственности. Вариативность окружающей среды может быть явно смоделирована путем изучения людей в широком диапазоне сред, хотя вывод о генетической дисперсии на основе фенотипической и экологической дисперсии может привести к недооценке наследуемости из-за проблемы захвата всего диапазона влияния окружающей среды, влияющего на признак. Другие методы расчета наследуемости используют данные из полногеномные ассоциации исследований для оценки влияния генетических факторов на признак, которое отражается скоростью и влиянием предположительно связанных генетических локусов (обычно однонуклеотидные полиморфизмы ) по признаку. Однако это может привести к недооценке наследуемости. Это несоответствие называется «отсутствующей наследуемостью» и отражает проблему точного моделирования как генетической, так и окружающей среды в моделях наследуемости.[23]
Когда доступна большая, сложная родословная или другой вышеупомянутый тип данных, наследуемость и другие количественные генетические параметры могут быть оценены следующим образом: ограниченная максимальная вероятность (REML) или Байесовские методы. В необработанные данные обычно будет иметь три или более точек данных для каждого индивидуума: код производителя, код матери и одно или несколько значений признаков. Различные значения признаков могут быть для разных признаков или для разных временных точек измерения.
Популярная в настоящее время методология основана на высокой степени уверенности в личности отца и матери; не принято относиться к отцовской идентичности вероятностно. Обычно это не проблема, поскольку методика редко применяется к диким популяциям (хотя она использовалась для нескольких популяций диких копытных и птиц), а производители неизменно известны с очень высокой степенью уверенности в программах разведения. Также существуют алгоритмы, учитывающие неопределенное отцовство.
Родословные можно просмотреть с помощью таких программ, как Pedigree Viewer. [1], и проанализированы с помощью таких программ, как ASReml, VCE [2], WOMBAT [3], MCMCglmm в среде R [4] или BLUPF90 семейство программ [5].
Родословные модели помогают распутать путаницу, такую как: обратная причинность, материнские эффекты такой как пренатальная среда, и смешение генетическое доминирование, общая среда и эффекты материнских генов.[24][9]
Геномная наследственность
Когда доступны данные о геноме всего генома и фенотипы из больших выборок населения, можно оценить отношения между людьми на основе их генотипов и использовать линейную смешанную модель для оценки дисперсии, объясняемой генетическими маркерами. Это дает оценку наследственности генома, основанную на дисперсии, зафиксированной в общих генетических вариантах.[4] Существует несколько методов, которые по-разному корректируют частоту аллелей и нарушение равновесия по сцеплению. В частности, метод под названием High-Definition Likelihood (HDL) может оценивать наследуемость генома, используя только сводную статистику GWAS,[5] упрощение включения большого размера выборки в различные метаанализы GWAS.
Ответ на выбор
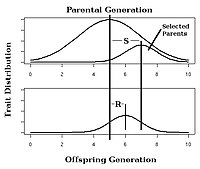
В селекция растений и животных, ожидаемый ответ на выбор признака с известной узкой наследуемостью можно оценить с помощью уравнение заводчика:[25]


В этом уравнении реакция на выбор (R) определяется как реализованная средняя разница между родительским поколением и следующим поколением, а дифференциал выбора (S) определяется как средняя разница между родительским поколением и выбранными родителями.[14]:1957[26]
Например, представьте, что селекционер участвует в проекте селекции с целью увеличения количества зерен на початок кукурузы. Ради аргументации предположим, что средний початок кукурузы в родительском поколении имеет 100 зерен. Предположим также, что выбранные родители производят кукурузу в среднем по 120 зерен на початок. Если h2 равно 0,5, то следующее поколение будет давать кукурузу в среднем 0,5 (120-100) = 10 дополнительных зерен на початок. Таким образом, общее количество зерен на початок кукурузы будет в среднем 110.
Наблюдение за реакцией на отбор в эксперименте с искусственным отбором позволит рассчитать реализованную наследуемость, как показано на рис. 4.
Обратите внимание, что наследуемость в приведенном выше уравнении равна отношению только если следуют генотип и шум окружающей среды Гауссовы распределения.

Споры
 | Эта секция может быть неуравновешенный к определенным точкам зрения. (Август 2016 г.) |
По оценке выдающихся критиков, таких как Стивен Роуз,[27] Джей Джозеф,[28] и Ричард Бенталл, сосредоточены в основном на оценках наследственности в поведенческие науки и социальные науки. Бенталл утверждал, что такие баллы наследуемости обычно вычисляются нелогично для получения высоких числовых баллов, что наследственность неверно интерпретируется как генетическое определение и что эта предполагаемая предвзятость отвлекает от других факторов, которые, по мнению исследователей, имеют более важное причинно-следственное значение, таких как жестокое обращение в детстве, вызывающее в дальнейшем психоз.[29][30] Оценки наследственности также по своей сути ограничены, потому что они не несут никакой информации о том, играют ли гены или окружающая среда большую роль в развитии изучаемого признака. Именно по этой причине, Дэвид Мур и Дэвид Шенк описывают термин «наследственность» в контексте генетики поведения как «... один из самых вводящих в заблуждение в истории науки» и утверждают, что он не имеет ценности, за исключением очень редких случаев.[31] При изучении сложных человеческих черт невозможно использовать анализ наследуемости для определения относительного вклада генов и окружающей среды, поскольку такие черты являются результатом взаимодействия множества причин.[32] Однако некоторые исследователи утверждают, что их можно разделить.[33]
Разногласия по поводу оценок наследственности в значительной степени связаны с их основанием в исследования близнецов. Скудный успех молекулярно-генетический исследования для подтверждения таких популяционно-генетический выводы исследований - это отсутствие наследственности проблема.[34] Эрик Туркхаймер утверждал, что новые молекулярные методы подтвердили традиционную интерпретацию исследований близнецов.[34] хотя по большей части остается неясным, как объяснить отношения между генами и поведением.[35] Согласно Туркхаймеру, и гены, и окружающая среда передаются по наследству, генетический вклад зависит от окружающей среды, а акцент на наследственности отвлекает от других важных факторов.[36] Однако в целом наследственность это широко применимая концепция.[9]
Смотрите также
использованная литература
- ^ Рэй Н., Вишер П. (2008). "Оценка наследуемости признака". Природное образование. 1 (1): 29. В архиве из оригинала от 2 августа 2015 г.. Получено 24 июля 2015.
- ^ Газзанига MS, Heatherton TF, Halpern DF. Психологическая наука (5-е изд.). Нью-Йорк. ISBN 978-0-393-26313-8. OCLC 908409996.
- ^ Ян Дж., Ли Ш., Годдард М.Э., Вишер П.М. (январь 2011 г.). «GCTA: инструмент для анализа комплексных признаков всего генома». Американский журнал генетики человека. 88 (1): 76–82. Дои:10.1016 / j.ajhg.2010.11.011. ЧВК 3014363. PMID 21167468.
- ^ а б Ян Дж., Зенг Дж., Годдард М.Э., Рэй Н.Р., Вишер П.М. (август 2017 г.). «Концепции, оценка и интерпретация наследуемости по SNP» (PDF). Природа Генетика. 49 (9): 1304–1310. Дои:10,1038 / нг.3941. PMID 28854176. S2CID 8790524. В архиве (PDF) из оригинала на 2020-10-05. Получено 2020-09-06.
- ^ а б Нин З, Павитан Й, Шен Х (июнь 2020 г.). «Вывод правдоподобия высокого разрешения генетических корреляций между комплексными чертами человека». Природа Генетика. 52 (8): 859–864. Дои:10.1038 / с41588-020-0653-у. HDL:10616/47311. PMID 32601477. S2CID 220260262.
- ^ Столтенберг С.Ф. (июнь 1997 г.). «Примирение с наследуемостью». Genetica. 99 (2–3): 89–96. Дои:10.1007 / BF02259512. HDL:2027.42/42804. PMID 9463077. S2CID 18212219.
- ^ Вальстен Д. (1994). «Разум наследственности» (PDF). Канадская психология. 35 (3): 244–260. Дои:10.1037/0708-5591.35.3.244. ISSN 1878-7304. В архиве (PDF) из оригинала на 2018-10-24. Получено 2019-12-05.
- ^ Maccoby EE (февраль 2000 г.). «Воспитание и его влияние на детей: генетика чтения и неправильного чтения». Ежегодный обзор психологии. 51 (1): 1–27. Дои:10.1146 / annurev.psych.51.1.1. PMID 10751963.
- ^ а б c Visscher PM, Hill WG, Wray NR (апрель 2008 г.). «Наследственность в эпоху геномики - концепции и заблуждения» (PDF). Обзоры природы. Генетика. 9 (4): 255–66. Дои:10.1038 / nrg2322. PMID 18319743. S2CID 690431. В архиве (PDF) из оригинала от 24.03.2016. Получено 2015-08-28.
- ^ Соус B, Matzel LD (январь 2018). «Парадокс интеллекта: наследственность и пластичность сосуществуют в скрытом взаимодействии генов и окружающей среды». Психологический бюллетень. 144 (1): 26–47. Дои:10.1037 / bul0000131. ЧВК 5754247. PMID 29083200.
- ^ Блок N (август 1995 г.). «Как наследственность вводит в заблуждение относительно расы». Познание. 56 (2): 99–128. Дои:10.1016 / 0010-0277 (95) 00678-р. PMID 7554794. S2CID 204981536.
- ^ Завещания C (2007). «Принципы популяционной генетики, 4-е издание» (PDF). Журнал наследственности (Рецензия на книгу). 98 (4): 382. Дои:10.1093 / jhered / esm035. В архиве (PDF) из оригинала на 2020-10-05. Получено 2018-04-20.
- обзор: Хартл Д.Л., Кларк А.Г. (2007). Принципы популяционной генетики. Сандерленд, Массачусетс: Sinauer and Associates. С. xv + 652. ISBN 978-0-87893-308-2.
- ^ Turkheimer E (октябрь 2000 г.). «Три закона генетики поведения и их значение» (PDF). Современные направления в психологической науке. 9 (5): 160–164. Дои:10.1111/1467-8721.00084. ISSN 0963-7214. S2CID 2861437. В архиве (PDF) из оригинала 19 октября 2013 г.. Получено 29 октября 2013.
- ^ а б c Кемпторн О. (1957). Введение в генетическую статистику (1-е изд.). Эймс, Айова: Университет штата Айова. Нажмите. OCLC 422371269.
- ^ Стивен Даунс и Лукас Мэтьюз. «Наследственность». Стэнфордская энциклопедия философии. Стэндфордский Университет. В архиве из оригинала на 25.02.2020. Получено 2020-02-20.
- ^ Зук О, Хехтер Э, Сюняев С.Р., Ландер Е.С. (январь 2012 г.). «Тайна отсутствия наследуемости: генетические взаимодействия создают фантомную наследуемость». Труды Национальной академии наук Соединенных Штатов Америки. 109 (4): 1193–8. Дои:10.1073 / pnas.1119675109. ЧВК 3268279. PMID 22223662.
- ^ Daw J, Guo G, Harris KM (июль 2015 г.). «Сеть заботы о природе: переоценка роли совместной среды в академической успеваемости и вербальном интеллекте». Исследования в области социальных наук. 52: 422–39. Дои:10.1016 / j.ssresearch.2015.02.011. ЧВК 4888873. PMID 26004471.
- ^ Кеттелл РБ (ноябрь 1960 г.). «Множественные абстрактные уравнения дисперсионного анализа и решения: для природоохранных исследований с непрерывными переменными». Психологический обзор. 67 (6): 353–72. Дои:10,1037 / ч0043487. PMID 13691636.
- ^ ДеФрис Дж. К., Фулкер Д. В. (сентябрь 1985 г.). «Множественный регрессионный анализ данных близнецов». Поведенческая генетика. 15 (5): 467–73. Дои:10.1007 / BF01066239. PMID 4074272. S2CID 1172312.
- ^ Falconer DS, Mackay TF (декабрь 1995 г.). Введение в количественную генетику (4-е изд.). Longman. ISBN 978-0582243026.
- ^ Gielen M, Lindsey PJ, Derom C, Smeets HJ, Souren NY, Paulussen AD, Derom R, Nijhuis JG (январь 2008 г.). «Моделирование генетических факторов и факторов окружающей среды для увеличения наследуемости и упрощения идентификации генов-кандидатов для определения веса при рождении: исследование близнецов». Поведенческая генетика. 38 (1): 44–54. Дои:10.1007 / s10519-007-9170-3. ЧВК 2226023. PMID 18157630.
- ^ Вальстен, Дуглас (март 1990). «Нечувствительность дисперсионного анализа к взаимодействию наследственности и среды» (PDF). Поведенческие науки и науки о мозге. 13 (1): 109–120. Дои:10.1017 / S0140525X00077797. ISSN 1469-1825.
- ^ Хекерман Д., Гурдасани Д., Кэди С., Помилла С., Карстенсен Т., Мартин Х., Экору К., Нсубуга Р. Н., Сеньёмо Г., Камали А., Калебу П., Видмер С., Сандху М. С. (июль 2016 г.). «Линейная смешанная модель для оценки наследуемости, которая явно учитывает изменения окружающей среды». Труды Национальной академии наук Соединенных Штатов Америки. 113 (27): 7377–82. Дои:10.1073 / pnas.1510497113. ЧВК 4941438. PMID 27382152.
- ^ Hill WG, Годдард ME, Visscher PM (февраль 2008 г.). Маккей Т.Ф., Годдард М.Э. (ред.). «Данные и теория указывают в основном на аддитивную генетическую дисперсию сложных признаков». PLOS Genetics. 4 (2): e1000008. Дои:10.1371 / journal.pgen.1000008. ЧВК 2265475. PMID 18454194.

- ^ Пломин Р., ДеФрис Дж. К., МакКлерн Г. Е., МакГаффин П. (2017). Поведенческая генетика: учебник (2-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-0-7167-2056-0.
- ^ Фалконер Д.С., Маккей Т.Ф. (1998). Введение в количественную генетику (4-е изд.). Эссекс: Лонгман. ISBN 978-0-582-24302-6.
- ^ Роза SP (Июнь 2006 г.). «Комментарий: оценки наследуемости - давно истек срок годности». Международный журнал эпидемиологии. 35 (3): 525–7. Дои:10.1093 / ije / dyl064. PMID 16645027.
- ^ Джозеф Дж (2004). «Глава 5». Генная иллюзия. Нью-Йорк: Алгора. п. 141. ISBN 978-1-898059-47-9. В архиве из оригинала на 19.07.2017. Получено 2016-04-02.
- ^ Бенталл РП (2009). Как лечить разум: действительно ли наше нынешнее лечение психических заболеваний хорошо?. Нью-Йорк: Издательство Нью-Йоркского университета. С. 123–127. ISBN 978-0-8147-8723-6. В архиве из оригинала на 2020-10-05. Получено 2016-04-02.
- ^ McGrath M (5 июля 2009 г.). «Лечение разума: обзор». Телеграф. В архиве из оригинала 4 июля 2012 г.. Получено 4 апреля 2018.
- ^ Мур Д.С., Шенк Д. (январь 2017 г.). «Заблуждение наследственности». Междисциплинарные обзоры Wiley: когнитивная наука. 8 (1–2): e1400. Дои:10.1002 / wcs.1400. PMID 27906501.
- ^ Фельдман М.В., Рамачандран С. (апрель 2018 г.). «Отсутствует по сравнению с чем? Возвращаясь к наследуемости, генам и культуре». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 373 (1743): 20170064. Дои:10.1098 / rstb.2017.0064. ЧВК 5812976. PMID 29440529.
... все сложные человеческие черты являются результатом сочетания причин. Если эти причины взаимодействуют, невозможно присвоить количественные значения доле признака, обусловленной каждой из них, так же как мы не можем сказать, какая часть площади прямоугольника обязана по отдельности каждому из двух его измерений. Таким образом, при анализе сложных фенотипов человека ... мы не можем найти «относительную важность генов и окружающей среды в определении фенотипа».
- ^ Треду, Гавань. «Природа и воспитание прямоугольников». (2019).
- ^ а б Туркхаймер Э (2011). "До сих пор отсутствует". Исследования в области человеческого развития. 8 (3–4): 227–241. Дои:10.1080/15427609.2011.625321. S2CID 14737438.
- ^ Туркхаймер Э (2015). «Генетическое предсказание». Отчет Центра Гастингса. 45 (5 Дополнение): S32–8. Дои:10.1002 / hast.496. PMID 26413946.
- ^ Джозеф Дж (2014). Проблема с исследованиями близнецов: переоценка исследований близнецов в социальных и поведенческих науках (PDF). Нью-Йорк: Рутледж. п. 81. ISBN 978-1-317-60590-4. В архиве (PDF) из оригинала от 04.04.2016. Получено 2016-04-02.
дальнейшее чтение
- Линч М., Уолш Б. (1998). Генетика и анализ количественных признаков. Сандерленд, Массачусетс: Sinauer Assoc. ISBN 978-0-87893-481-2.
- Джонсон В., Пенке Л., Spinath FM (2011). «Понимание наследственности: что это такое, а что нет». Европейский журнал личности. 25 (4): 287–294. Дои:10.1002 / пер.835. ISSN 0890-2070.
внешние ссылки
- Стэнфордская энциклопедия философии запись о наследственности и наследственности
- Веб-сайт Quantitative Genetics Resources, в том числе двухтомник Линча и Уолша. Бесплатный доступ
| Концепции количественной генетики | |
|---|---|
| Похожие темы | |
| |
| Лысенковцы | |
|---|---|
| Диссиденты | |
| Концепции | |
| Советская политика | |